|
The INTERNATIONAL JOURNAL of APPLIED RESEARCH In Veterinary Medicine |
 |
| Current Issue |
| Previous Issues |
| Reprint Information |
| Back to The International Journal of Applied Research in Veterinary Medicine |
Gene Expression
in the Early Phase of Murine Influenza Pneumonia Determined by cDNA
Expression Array Technique
Shinya Sakai
Hiroshi Ochiai
Naoki Mantani*
Toshiaki Kogure*
Yutaka Shimada§
Katsutoshi Terasawa§
Department of Kampo Diagnostics, Institute of Natural Medicine, Toyama Medical and Pharmaceutical University, Toyama, Japan.
Department of Human Science,
Toyama Medical and Pharmaceutical University, Toyama, Japan.
§Department of Japanese Oriental (Kampo) Medicine, Faculty of Medicine, Toyama Medical and Pharmaceutical University, Toyama, Japan.
*Department
of Integrated Japanese Oriental Medicine, School of Medicine, Gunma
University, Gunma, Japan.
KEY WORDS: influenza, cDNA array, chemokine, cytokine, murine
Abstract
To assess the influence of influenza virus infection on gene expression in mice, mRNA levels in lung and tracheal tissue 48 hours after infection were investigated by cDNA array analysis. We examined the expression of 65 growth factors, cytokines, and chemokines mRNAs. The expression of 6 mRNAs, small inducible cytokine B subfamily member (SCYB) 10 , SCYB 9, small inducible cytokine A2, wingless-related MMTV integration site 8b protein, wingless-related MMTV integration site 1 protein precursor and fibroblast inducible secreted protein, were upregulated. Vascular endothelial growth factor was downregulated. Furthermore, we examined the expression of 50 receptors mRNAs of growth factors, cytokines, and chemokines. As a result, the expression of 5 mRNAs that is fibroblast growth factor receptor 1, fibroblast growth factor receptor 4, interleukin (IL)-10 receptor alpha, colony stimulating factor 2 receptor, beta 1, and IL-9 receptor were upregulated. Tumor necrosis factor receptor subfamily member 9 was downregulated. Although further studies are required, these data suggest that these cytokines play an important role in influenza virus infection especially in the phase before specific immunity.
Introduction
Despite vaccines and antiviral substances, influenza still causes significant morbidity and mortality worldwide. Better understanding of the molecular mechanisms of influenza virus pathogenesis and host immune responses is required for the development of more efficient means of preventing and treating of influenza. Many features of the antiviral immune response have been elucidated using influenza viruses adapted to mice. Intranasal infection of mice results in cellular infiltration in the respiratory tract, with interstitial pneumonia similar to that observed in severe human influenza.
Although 15 to 20 genes have been characterized as showing increased expression after infection of the respiratory tract with influenza virus, these genes probably represent only a small fraction of all the genes that are induced under these circumstances. Little is known about the genes involved in the inflammatory response with influenza virus. In particular, little information has been gathered about the early phase of this response, before the appearance of specific cytotoxic T cells. During this phase, influenza virus interacts with cells on the luminal side of the airways and alveoli to induce the release of immunoactive mediators, thus initiating a cascade of events resulting in the eventual elimination of the virus.
Recently, techniques have become available that allow characterization of the mRNA expression status of the large number of genes.1,2 In this study we examined mRNA expression in murine influenza pneumonia using cDNA array technique especially focusing on cytokines and their receptors.
Materials and Methods
Preparation of Virus
The lung-adapted strain of influenza A/PR/8/34 (PR8) virus (H1N1 subtype) was propagated in the chorioallantoic cavity of 10-day-old embryonated hen eggs for 48 hours at 35ºC. The chorioallantoic fluid was collected and stored in small portions at 80˚C after centrifugation at 1,000 ¥ g for 10 minutes. The virus titer of the chorioallantoic fluid was 1.9 ¥ 108 plaque forming units (PFU) as determined on Mardin-Darby canine kidney cells as described previously.3
Virus Infection of Mice and
Sample Preparation
Outbred specific-pathogen-free ICR female 4-week-old mice (body weight, approximately 17 g) were obtained from SLC Company (Hamamatsu, Japan) and were infected by intranasal inoculation of a virus solution containing 1.0 ¥ 104 PFU/25 µL (five 50% lethal doses of virus) under ether anesthesia. The mice were killed 48 hours after infection, and the tracheas and lungs were removed. The resected trachea and lungs were frozen by liquid nitrogen immediately after resection.
cDNA expression array: To determine gene expression, the membrane-based microtechnique with an Atlas cDNA expression array (mouse 1.2 array II; Clontech) was performed in accordance with the manual provided. The array included 1,176 mouse cDNA and 9 housekeeping control cDNA and negative controls immobilized on a nylon membrane. The cDNA on a membrane is divided into 23 categories: (1) 46 cDNA for cell surface antigens; (2) 65 cDNAs for growth factors, cytokines, and chemokines; (3) 25 cDNA for hormones; (4) 23 cDNA for RNA processing, turnover, and transport; (5) 66 cDNA for transcription factors and DNA-binding proteins; (6) 7 cDNA for cell cycle regulators; (7) 45 cDNA for cell adhesion receptors and proteins, (8) 89 cDNA for extracellular transporters; (9) 7 cDNA for oncogenes and tumor suppressors, (10) 14 cDNA for stress response proteins; (11) 24 cDNA for extracellular matrix proteins; (12) 42 cDNA for trafficking and targeting protein; (13) 187 cDNA for metabolic pathways, (14) 3 cDNA for post-translational modification and folding; (15) 14 cDNA for translation; (16) 3 cDNA for apoptosis-associated proteins; (17) 164 cDNA for receptors; (18) 28 cDNA for extracellular cell signaling and communication; (19) 69 cDNA for modulators, effectors, and intracellular transducers; (20) 10 cDNA for protein turnover; (21) 21 cDNA for cytoskelton and motility proteins; (22) 13 cDNA for DNA synthesis, repair, and recombination proteins; (23) 214 cDNA for other, and 9 cDNA for housekeeping genes.
The purified RNA,
which was analyzed for genomic DNA contamination by PCR with primers
specific for ß-actin, was processed with the gene-specific CDS primer
mix (Clontech), deoxynucleoside triphosphate, [32P]dATP, and reverse
transcriptase for preparation of cDNA. The 32P-labeled cDNA was purified
through a Chroma Spin-200 column (Clontech). The labeled cDNA in a
solution of ExpressHyb (Clontech) with heat-denatured, sheared-salmon-test
DNA was then hybridized overnight to an Atlas array membrane at 68˚C.
The membrane was washed in 2XSSC (1XSSC is 0.15 M NaCl plus 0.015
M sodium citrate) with 1% sodium dodecyl sulfate, 0.1XSSC with 0.5%
sodium dodecyl sulfate, and 2XSSC, sequentially, then exposed to PhosphorImager
(Storm 860; Molecular Dynamics, Sunnyvale, CA, U.S.A.). Results of
the gene expression were analyzed by computer with Atlas image software
(Clontech).
Statistic Analysis
Comparison of results of the gene expression between noninfected control mice and infected mice was analyzed by Students t-test.
Results
Table 1 shows the cDNA for growth factors, cytokines, and chemokines that were tested in this study. Of 65 mRNA examined, 6 mRNA were upregulated and one mRNA was downregulated at 48 hours after infection (Figure 1). Gene expression levels (spot intensity) of small inducible cytokine B subfamily member (SCYB) 10, SCYB9, small inducible cytokine A2, wingless-related MMTV integration site 8b protein, wingless-related MMTV integration site 1 protein precursor and fibroblast inducible secreted protein before infection were 1.0 ± 0.0, 1.0 ± 0.6, 1.0 ± 0.3, 2.0 ± 0.3, 1.0 ± 0.0, 7.0 ± 1.2, respectively. At 48 hours after infection, gene expression levels of small inducible cytokine B subfamily member (SCYB) 10, SCYB9, small inducible cytokine A2, wingless-related MMTV integration site 8b protein, wingless-related MMTV integration site 1 protein precursor and fibroblast inducible secreted protein before infection were 93.0 ± 21.4, 16.0 ± 3.8, 20.0 ± 3.6, 14.0 ± 0.9, 18.0 ± 4.4, 12.0 ± 1.2, respectively. Gene expression level of vascular endothelial growth factor before infection was 10.0 ± 3.7. However, the level was 5.0 ± 1.0 at 48 hours after infection.
Table 2 shows the
cDNA for growth factors, cytokines, and chemokines receptors that
were tested in this study. Of 50, 5 mRNA were upregulated, and one
mRNA was downregulated at 48 hours after infection (Figure 2). Gene
expression levels (spot intensity) of fibroblast growth factor receptor
1, fibroblast growth factor receptor 4, interleukin (IL)-10 receptor
alpha, colony stimulating factor 2 receptor, beta 1 and IL-9 receptor
before infection were 5.0 ± 1.3, 2.0 ± 1.0, 7.0 ± 1.5, 0.0 ± 0.3,
2.0 ± 0.6, respectively. Gene expression levels of tumor necrosis
factor (TNF) receptor subfamily member 9 before and 48 hours after
infection were 8.0 ± 3.2 and 3.0 ± 0.9, respectively.
Discussion
In this study, we examined the expression of 65
growth factors, cytokines, and chemokines mRNA. The expression of
6 mRNA, SCYB 10, SCYB9, small inducible cytokine A2, wingless-related
MMTV integration site 8b protein, wingless-related MMTV integration
site 1 protein precursor, and fibroblast-inducible secreted protein,
were upregulated. Vascular endothelial growth factor was downregulated.
Although how these cytokines work in the influenza infection site
is not clear, this is first time to show these cytokines and related
proteins mRNA are up regulated in infection sites.
We also examined
the expression of 50 mRNA of growth factors receptors, cytokines receptors,
and chemokines receptors. As a result, the expression of 5 mRNA, fibroblast
growth factor receptor 1, fibroblast growth factor receptor 4, IL-10
receptor alpha, colony stimulating factor 2 receptor, beta 1, and
IL-9 receptor, were upregulated. TNF-receptor subfamily member 9 was
downregulated.
Researchers have
shown that proinflammatory cytokines such as IL-1, IL-6, and TNF are
produced by influenza virus infection.4,5 However, in this study,
we did not observe the upregulation of IL-1 and TNF mRNA. Our observation
was 48 hours after infection. It is likely that the time point of
observation was the phase between innate immunity and specific immunity.
Therefore, it is interesting that the mRNA expression of these cytokines
and receptors were upregulated or downregulated. Although further
studies are required, these data suggest that these cytokines play
an important role in influenza virus infection especially during the
phase between innate immunity and specific immunity.
References
1. Coombes BK, Mahony JB: cDNA array analysis of altered gene expression in human endothelial cells in response to Chlamydia pneumoniae infection. Infection & Immunity 69:1420-1427, 2001.
2. Eckmann L, Smith JR, Housley MP, et al: Analysis by high density cDNA arrays of altered gene expression in human intestinal epithelial cells in response to infection with the invasive enteric bacteria Salmonella. J Biol Chem 275:14084-14094, 2001.
3. Ochiai H, Ikesue A, Kurokawa M, et al: Enhanced production of rat interleukin-8 by in vitro and in vivo infections with influenza A NWS virus. J Virol 67:6811-6814, 1993.
4. Julkunen I, Sareneva T, Pirhonen J, et al: Molecular pathogenesis of influenza A virus infection and virus-induced regulation of cytokine gene expression. Cytokine and Growth Factor Reviews 12:171-180, 2001.
5.
Stark GK, Williams BR, Silverman RH, Schreiber RD: How cells
respond to interferons. Annu Rev Biochem 67:227-264, 1998.
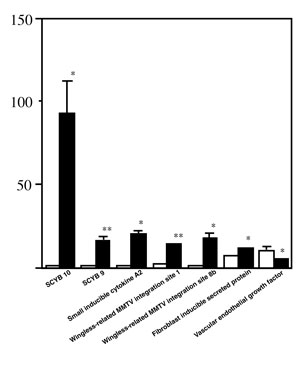
Figure 1. mRNA
expression levels for growth factors, cytokines, and chemokines in
lung and trachea tissue of influenza infected mice. Results represented
are means plus standard error from three independent experiments.
Open column, before infection; closed column, 48-h after infection
of influenza virus. *P < 0.05; **P < 0.01 compared with noninfected
controls analyzed by Students t-test.

Figure
2. mRNA expression levels
for growth factors, cytokines, and chemokine receptors in lung and
trachea tissue of influenza infected mice. Results represented are
means plus standard error from three independent experiments. Open
column, before infection; closed column, 48 hours after infection
of influenza virus. *P < 0.05; **P < 0.01 compared with noninfected
controls analyzed using Students t-test.
Table
2. Receptor
mRNAs of Growth Factors, Cytokines, and Chemokines
activin A receptor, type 1B kit oncogene
activin A receptor, type II-like 1 nerve growth factor receptor
Burkitt lymphoma receptor 1 platelet derived growth factor receptor, alpha polypeptide
chemokine (C-C) receptor 2 platelet derived growth factor receptor, beta polypeptide
chemokine (C-C) receptor 4 macrophage stimulating 1 receptor
(c-met-related tyrosine kinase)
chemokine (C-C) receptor 5 tumor necrosis factor receptor superfamily, member 11a
chemokine (C-C) receptor 7 tumor necrosis factor receptor superfamily, member 1b
D6 beta-chemokine receptor tumor necrosis factor receptor superfamily, member 9**
chemokine (C-X3-C) receptor 1 tumor necrosis factor receptor superfamily, member 1a
chemokine (C-X-C) receptor 2 interleukin 10 receptor, alpha*
chemokine (C-X-C) receptor 3 interleukin 10 receptor, beta
colony stimulating factor 1 receptor interleukin 11 receptor, alpha chain 1
colony
stimulating factor 3 receptor
interleukin 12 receptor, beta 2
(granulocyte)
endothelial-specific receptor tyrosine kinase interleukin 13 receptor, alpha 1
erythropoietin receptor interleukin 13 receptor, alpha 2
fibroblast growth factor receptor 1* interleukin 15 receptor, alpha chain
fibroblast growth factor receptor 2 interleukin 17 receptor
fibroblast growth factor receptor 4* interleukin 2 receptor, alpha chain
FMS-like tyrosine kinase 1 interleukin 2 receptor, beta chain
FMS-like tyrosine kinase 3 interleukin 3 receptor, alpha chain
FMS-like
tyrosine kinase 4
colony stimulating factor 2 receptor,
beta 1, low-affinity (granulocyte-macrophage)*
glial cell line derived neurotrophic
factor interleukin 5 receptor, alpha
family receptor alpha 2
glial cell line derived neurotrophic
factor interleukin 6 receptor, alpha
amily receptor alpha 1
insulin-like growth factor I receptor interleukin 7 receptor
kinase insert domain protein receptor interleukin 9 receptor*
*upregulated,
**downregulated
ISSN# 1542-2666